HP1-Driven Micro-Phase Separation of Heterochromatin-Like Domains/Complexes.
IF 3.6
Q2 GENETICS & HEREDITY
Epigenetics Insights
Pub Date : 2022-07-05
eCollection Date: 2022-01-01
DOI:10.1177/25168657221109766
引用次数: 2
Abstract
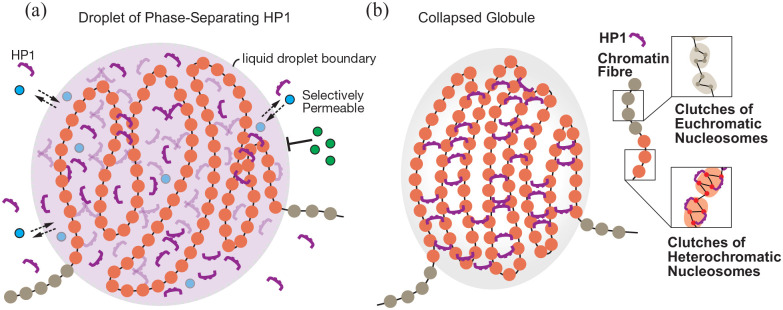
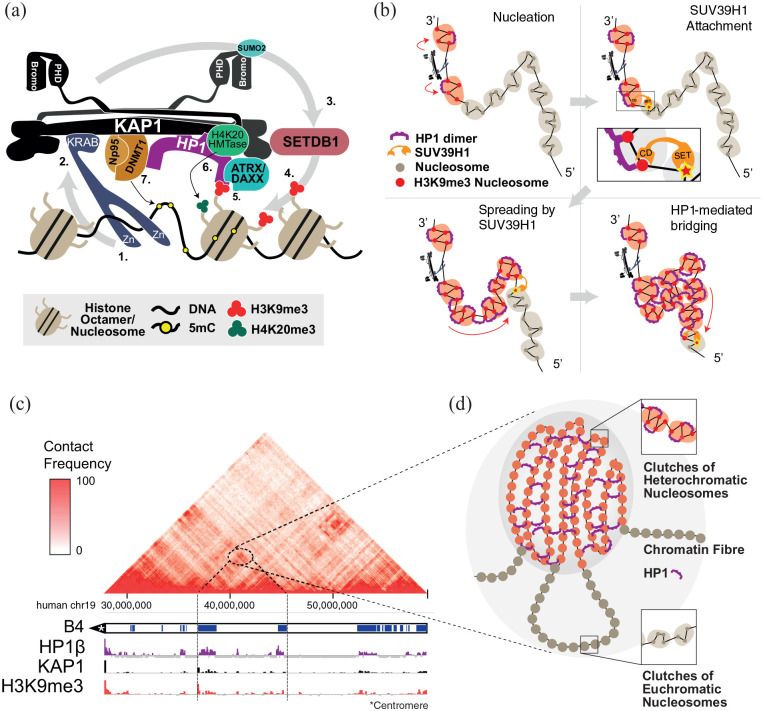
Creative Commons Non Commercial CC BY-NC: This article is distributed under the terms of the Creative Commons Attribution-NonCommercial 4.0 License (https://creativecommons.org/licenses/by-nc/4.0/) which permits non-commercial use, reproduction and distribution of the work without further permission provided the original work is attributed as specified on the SAGE and Open Access pages (https://us.sagepub.com/en-us/nam/open-access-at-sage). Cytologically visible constitutive heterochromatin found at the centromeric and telomeric regions of chromosomes represents the largest differentiated chromatin compartment in eukaryotic nuclei.1 Outside constitutive heterochromatin, heterochromatin-like domains (HLD, >.1 Mb) and complexes (HLC, <.1 Mb) are found along the chromosome arms.2 HLD/Cs are present in eukaryotic genomes as divergent as fission yeast and human, with expansions in size and number in mammals. In human, there are ~163 to 859 HLDs and ~18 853 to 32 292 HLCs, depending on cell type.2 Some of the largest HLDs are the KRAB-ZNF HLDs on chromosome 19. Notably, contacts among these large HLDs emerge as the heterochromatic B4 sub-compartment in Hi-C maps.3 Both cytologically visible constitutive heterochromatin and HLD/Cs are associated with the di/tri-methyl modification of lysine 9 on histone H3 (H3K9me2/3) and the HP1-class of chromodomain (CD) proteins.1,2 The HP1 CD binds to H3K9me2/3 and HP1 proteins dimerise through the chromo shadow domain (CSD), creating ‘bridges’ between 2 H3K9methylated molecules in separate nucleosomes.4 How the HP1 and H3K9me2/3-marked chromatin are involved in partitioning the genome into cytologically visible constitutive heterochromatin and euchromatin as well as into the heterochromatic B-type and euchromatic A-type compartments has been the subject of intense research and led to the notion that the partitioning observed is driven by the same physiochemical process, namely phase separation. However, recent work shows that HP1-driven liquid-liquid phase separation (LLPS) is unlikely to play a major role in the formation and establishment of constitutive heterochromatin. By contrast, HP1 proteins appear to be key players in micro-phase separation and segregation of HLD/ Cs that generate heterochromatic B-type compartments.

hp1驱动的异染色质样结构域/复合物的微相分离。
本文章由计算机程序翻译,如有差异,请以英文原文为准。
求助全文
约1分钟内获得全文
求助全文

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


