{"title":"Correction to “cIAP-2 and Survivin Contribute to Cytokine-Mediated Delayed Eosinophil Apoptosis”","authors":"","doi":"10.1002/eji.202570063","DOIUrl":null,"url":null,"abstract":"<p>E. M. Vassina, S. Yousefi, D. Simon, C. Zwicky, S. Conus, and H.-U. Simon, “CIAP-2 and Survivin Contribute to Cytokine-Mediated Delayed Eosinophil Apoptosis,” <i>European Journal of Immunology</i> 36, no. 7 (2006): 1975–1984, https://doi.org/10.1002/eji.200635943.</p><p>Concerns were raised by a third party regarding duplicated image panels within Figure 2A, between the cIAP-1 and cIAP-2 normal neutrophil subpanels. The authors acknowledged the image compilation error, and as the original raw data were no longer available, they repeated the experiment. The new data confirmed that the corresponding experimental results and the overall conclusions of the paper remain unaffected. The corrected panels of Figure 2A, the full-length immunoblots, and the updated methods are below. The authors apologize for this mistake.</p><p><b>Corrected image panels of Figure</b> 2A</p><p>Lack of expression of cIAP-1 and cIAP-2 in freshly purified normal human neutrophils compared to THP-1 and HL-60 cell lines. Immunoblot analysis was performed to assess protein expression. No detectable levels of cIAP-1 or cIAP-2 were observed in normal blood neutrophils. Lysates from THP-1 and HL-60 cells served as controls. Membranes were re-probed with anti-GAPDH monoclonal antibody to confirm equal protein loading.</p><p><b>Full-length immunoblots of the repeated experiments</b></p><p>Full-length immunoblots are shown for cIAP-1, cIAP-2, and GAPDH protein expression. Each lane was loaded with 50 µg of total cell lysate derived from either human blood neutrophils or the control cell lines THP-1 and HL-60. GAPDH served as a loading control.</p><p>Peripheral blood neutrophils from control individuals were purified as described [<span>1</span>] and were >98% pure. For control experiments, the human promyelocytic leukemia cell line HL-60 clone 15 and the human monocytic leukemia cell line THP-1 (both from ATCC, Manassas, VA, USA) were used.</p><p>Cell-free extracts and immunoblotting were performed as previously described [<span>2</span>]. Briefly, cell pellets were resuspended in lysis buffer (50 mM Tris-HCl [pH 7.4], 150 mM NaCl, 10% glycerol, 1% Triton X-100, 1% NP-40, 2 mM EDTA, 2.5 mM MgCl₂, 2.5 mM NaF, 10 mM sodium pyrophosphate, and 200 µM sodium orthovanadate), freshly supplemented with protease inhibitor cocktail (Sigma-Aldrich), 1 mM PMSF (Sigma-Aldrich), and 1 × PhosSTOP phosphatase inhibitor cocktail (Roche). Cells were collected, washed with PBS, and lysed on ice for 20 min. Lysates were clarified by high-speed centrifugation (13,000 rpm, 15 min, 4 °C). Protein concentrations were determined using the Pierce BCA protein assay kit (Thermo Fisher Scientific).</p><p>Equal amounts of protein (50 µg) were denatured and separated on 12% SERVAGel TG PRiME gels (SERVA Electrophoresis, Heidelberg, Germany), followed by transfer onto Immobilon-P PVDF membranes (Merck Millipore). Membranes were blocked with 5% nonfat dry milk in TBST (20 mM Tris-HCl, 150 mM NaCl, 0.1% Tween 20, pH 7.6) for 1 h at room temperature, and incubated overnight at 4 °C with primary antibodies.</p><p>The following antibodies were used: monoclonal rabbit anti-human cIAP-1 (clone D5G9; 1:1000 dilution) and cIAP-2 (clone 58C7; 1:500 dilution) (both from Cell Signaling Technology, distributed by BioConcept Ltd., Allschwil, Switzerland) and monoclonal mouse anti-human GAPDH (clone 6C5; 1:2000 dilution; Merck Millipore, Darmstadt, Germany). After washing, membranes were incubated with horseradish peroxidase (HRP)-conjugated sheep anti-mouse secondary antibody (1:5000 dilution) for 1 h at room temperature. Signals were developed using the Immobilon Forte Western HRP substrate (Merck Millipore) and visualized using the Odyssey Fc Imaging System (LI-COR Biosciences, Lincoln, USA).</p>","PeriodicalId":165,"journal":{"name":"European Journal of Immunology","volume":"55 7","pages":""},"PeriodicalIF":3.7000,"publicationDate":"2025-07-09","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/eji.202570063","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"European Journal of Immunology","FirstCategoryId":"3","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/eji.202570063","RegionNum":3,"RegionCategory":"医学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q2","JCRName":"IMMUNOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
E. M. Vassina, S. Yousefi, D. Simon, C. Zwicky, S. Conus, and H.-U. Simon, “CIAP-2 and Survivin Contribute to Cytokine-Mediated Delayed Eosinophil Apoptosis,” European Journal of Immunology 36, no. 7 (2006): 1975–1984, https://doi.org/10.1002/eji.200635943.
Concerns were raised by a third party regarding duplicated image panels within Figure 2A, between the cIAP-1 and cIAP-2 normal neutrophil subpanels. The authors acknowledged the image compilation error, and as the original raw data were no longer available, they repeated the experiment. The new data confirmed that the corresponding experimental results and the overall conclusions of the paper remain unaffected. The corrected panels of Figure 2A, the full-length immunoblots, and the updated methods are below. The authors apologize for this mistake.
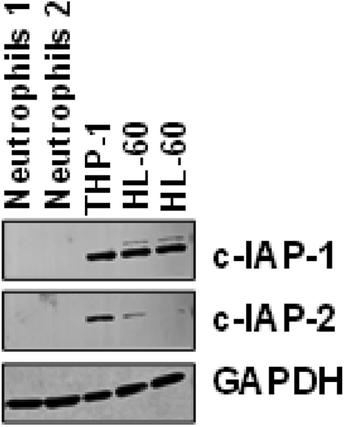
Corrected image panels of Figure 2A
Lack of expression of cIAP-1 and cIAP-2 in freshly purified normal human neutrophils compared to THP-1 and HL-60 cell lines. Immunoblot analysis was performed to assess protein expression. No detectable levels of cIAP-1 or cIAP-2 were observed in normal blood neutrophils. Lysates from THP-1 and HL-60 cells served as controls. Membranes were re-probed with anti-GAPDH monoclonal antibody to confirm equal protein loading.
Full-length immunoblots of the repeated experiments
Full-length immunoblots are shown for cIAP-1, cIAP-2, and GAPDH protein expression. Each lane was loaded with 50 µg of total cell lysate derived from either human blood neutrophils or the control cell lines THP-1 and HL-60. GAPDH served as a loading control.
Peripheral blood neutrophils from control individuals were purified as described [1] and were >98% pure. For control experiments, the human promyelocytic leukemia cell line HL-60 clone 15 and the human monocytic leukemia cell line THP-1 (both from ATCC, Manassas, VA, USA) were used.
Cell-free extracts and immunoblotting were performed as previously described [2]. Briefly, cell pellets were resuspended in lysis buffer (50 mM Tris-HCl [pH 7.4], 150 mM NaCl, 10% glycerol, 1% Triton X-100, 1% NP-40, 2 mM EDTA, 2.5 mM MgCl₂, 2.5 mM NaF, 10 mM sodium pyrophosphate, and 200 µM sodium orthovanadate), freshly supplemented with protease inhibitor cocktail (Sigma-Aldrich), 1 mM PMSF (Sigma-Aldrich), and 1 × PhosSTOP phosphatase inhibitor cocktail (Roche). Cells were collected, washed with PBS, and lysed on ice for 20 min. Lysates were clarified by high-speed centrifugation (13,000 rpm, 15 min, 4 °C). Protein concentrations were determined using the Pierce BCA protein assay kit (Thermo Fisher Scientific).
Equal amounts of protein (50 µg) were denatured and separated on 12% SERVAGel TG PRiME gels (SERVA Electrophoresis, Heidelberg, Germany), followed by transfer onto Immobilon-P PVDF membranes (Merck Millipore). Membranes were blocked with 5% nonfat dry milk in TBST (20 mM Tris-HCl, 150 mM NaCl, 0.1% Tween 20, pH 7.6) for 1 h at room temperature, and incubated overnight at 4 °C with primary antibodies.
The following antibodies were used: monoclonal rabbit anti-human cIAP-1 (clone D5G9; 1:1000 dilution) and cIAP-2 (clone 58C7; 1:500 dilution) (both from Cell Signaling Technology, distributed by BioConcept Ltd., Allschwil, Switzerland) and monoclonal mouse anti-human GAPDH (clone 6C5; 1:2000 dilution; Merck Millipore, Darmstadt, Germany). After washing, membranes were incubated with horseradish peroxidase (HRP)-conjugated sheep anti-mouse secondary antibody (1:5000 dilution) for 1 h at room temperature. Signals were developed using the Immobilon Forte Western HRP substrate (Merck Millipore) and visualized using the Odyssey Fc Imaging System (LI-COR Biosciences, Lincoln, USA).
期刊介绍:
The European Journal of Immunology (EJI) is an official journal of EFIS. Established in 1971, EJI continues to serve the needs of the global immunology community covering basic, translational and clinical research, ranging from adaptive and innate immunity through to vaccines and immunotherapy, cancer, autoimmunity, allergy and more. Mechanistic insights and thought-provoking immunological findings are of interest, as are studies using the latest omics technologies. We offer fast track review for competitive situations, including recently scooped papers, format free submission, transparent and fair peer review and more as detailed in our policies.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


