Aiko Iwata-Otsubo, Cara M Skraban, Atsunori Yoshimura, Toyonori Sakata, Cesar Augusto P Alves, Sarah K Fiordaliso, Yukiko Kuroda, Jaime Vengoechea, Angela Grochowsky, Paige Ernste, Lauren Lulis, Addie Nesbitt, Ahmad Abou Tayoun, Christopher Gray, Meghan C Towne, Kelly Radtke, Elizabeth A Normand, Lindsay Rhodes, Christoph Seiler, Katsuhiko Shirahige, Kosuke Izumi
{"title":"Biallelic variants in GTF3C5, a regulator of RNA polymerase III-mediated transcription, cause a multisystem developmental disorder.","authors":"Aiko Iwata-Otsubo, Cara M Skraban, Atsunori Yoshimura, Toyonori Sakata, Cesar Augusto P Alves, Sarah K Fiordaliso, Yukiko Kuroda, Jaime Vengoechea, Angela Grochowsky, Paige Ernste, Lauren Lulis, Addie Nesbitt, Ahmad Abou Tayoun, Christopher Gray, Meghan C Towne, Kelly Radtke, Elizabeth A Normand, Lindsay Rhodes, Christoph Seiler, Katsuhiko Shirahige, Kosuke Izumi","doi":"10.1007/s00439-024-02656-3","DOIUrl":null,"url":null,"abstract":"<p><p>General transcription factor IIIC subunit 5 (GTF3C5) encodes transcription factor IIIC63 (TFIIIC63). It binds to DNA to recruit another transcription factor, TFIIIB, and RNA polymerase III (Pol III) to mediate the transcription of small noncoding RNAs, such as tRNAs. Here, we report four individuals from three families presenting with a multisystem developmental disorder phenotype with biallelic variants in GTF3C5. The overlapping features include growth retardation, developmental delay, intellectual disability, dental anomalies, cerebellar malformations, delayed bone age, skeletal anomalies, and facial dysmorphism. Using lymphoblastoid cell lines (LCLs) from two affected individuals, we observed a reduction in TFIIIC63 protein levels compared to control LCLs. Genome binding of TFIIIC63 protein is also reduced in LCL from one of the affected individuals. Additionally, approximately 40% of Pol III binding regions exhibited reduction in the level of Pol III occupancy in the mutant genome relative to the control, while approximately 54% of target regions showed comparable levels of Pol III occupancy between the two, indicating partial impairment of Pol III occupancy in the mutant genome. Yeasts with subject-specific variants showed temperature sensitivity and impaired growth, supporting the notion that the identified variants have deleterious effects. gtf3c5 mutant zebrafish showed developmental defects, including a smaller body, head, and eyes. Taken together, our data show that GTF3C5 plays an important role in embryonic development, and that biallelic variants in this gene cause a multisystem developmental disorder. Our study adds GTF3C5-related disorder to the growing list of genetic disorders associated with Pol III transcription machinery.</p>","PeriodicalId":13175,"journal":{"name":"Human Genetics","volume":" ","pages":"437-453"},"PeriodicalIF":3.8000,"publicationDate":"2024-03-01","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Human Genetics","FirstCategoryId":"99","ListUrlMain":"https://doi.org/10.1007/s00439-024-02656-3","RegionNum":2,"RegionCategory":"生物学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"2024/3/23 0:00:00","PubModel":"Epub","JCR":"Q2","JCRName":"GENETICS & HEREDITY","Score":null,"Total":0}
引用次数: 0
Abstract
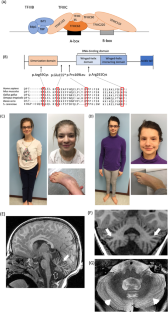
General transcription factor IIIC subunit 5 (GTF3C5) encodes transcription factor IIIC63 (TFIIIC63). It binds to DNA to recruit another transcription factor, TFIIIB, and RNA polymerase III (Pol III) to mediate the transcription of small noncoding RNAs, such as tRNAs. Here, we report four individuals from three families presenting with a multisystem developmental disorder phenotype with biallelic variants in GTF3C5. The overlapping features include growth retardation, developmental delay, intellectual disability, dental anomalies, cerebellar malformations, delayed bone age, skeletal anomalies, and facial dysmorphism. Using lymphoblastoid cell lines (LCLs) from two affected individuals, we observed a reduction in TFIIIC63 protein levels compared to control LCLs. Genome binding of TFIIIC63 protein is also reduced in LCL from one of the affected individuals. Additionally, approximately 40% of Pol III binding regions exhibited reduction in the level of Pol III occupancy in the mutant genome relative to the control, while approximately 54% of target regions showed comparable levels of Pol III occupancy between the two, indicating partial impairment of Pol III occupancy in the mutant genome. Yeasts with subject-specific variants showed temperature sensitivity and impaired growth, supporting the notion that the identified variants have deleterious effects. gtf3c5 mutant zebrafish showed developmental defects, including a smaller body, head, and eyes. Taken together, our data show that GTF3C5 plays an important role in embryonic development, and that biallelic variants in this gene cause a multisystem developmental disorder. Our study adds GTF3C5-related disorder to the growing list of genetic disorders associated with Pol III transcription machinery.
通用转录因子 IIIC 亚基 5(GTF3C5)编码转录因子 IIIC63(TFIIIC63)。它与 DNA 结合,招募另一个转录因子 TFIIIB 和 RNA 聚合酶 III(Pol III),介导小的非编码 RNA(如 tRNA)的转录。在此,我们报告了来自三个家庭的四名患者,他们的多系统发育障碍表型与 GTF3C5 双重变异有关。其重叠特征包括生长迟缓、发育迟缓、智力障碍、牙齿异常、小脑畸形、骨龄延迟、骨骼异常和面部畸形。通过使用来自两个受影响个体的淋巴母细胞系(LCLs),我们观察到与对照 LCLs 相比,TFIIIC63 蛋白水平有所降低。在其中一个受影响个体的 LCL 中,TFIIIC63 蛋白的基因组结合也减少了。此外,与对照组相比,突变体基因组中约有 40% 的 Pol III 结合区的 Pol III 占有率水平降低,而约有 54% 的目标区的 Pol III 占有率水平与对照组相当,这表明突变体基因组中 Pol III 占有率部分受损。gtf3c5突变体斑马鱼表现出发育缺陷,包括身体、头部和眼睛较小。综上所述,我们的数据表明 GTF3C5 在胚胎发育中扮演着重要角色,而且该基因的双倍变体会导致多系统发育障碍。我们的研究为与 Pol III 转录机制相关的遗传性疾病增加了 GTF3C5 相关疾病。
期刊介绍:
Human Genetics is a monthly journal publishing original and timely articles on all aspects of human genetics. The Journal particularly welcomes articles in the areas of Behavioral genetics, Bioinformatics, Cancer genetics and genomics, Cytogenetics, Developmental genetics, Disease association studies, Dysmorphology, ELSI (ethical, legal and social issues), Evolutionary genetics, Gene expression, Gene structure and organization, Genetics of complex diseases and epistatic interactions, Genetic epidemiology, Genome biology, Genome structure and organization, Genotype-phenotype relationships, Human Genomics, Immunogenetics and genomics, Linkage analysis and genetic mapping, Methods in Statistical Genetics, Molecular diagnostics, Mutation detection and analysis, Neurogenetics, Physical mapping and Population Genetics. Articles reporting animal models relevant to human biology or disease are also welcome. Preference will be given to those articles which address clinically relevant questions or which provide new insights into human biology.
Unless reporting entirely novel and unusual aspects of a topic, clinical case reports, cytogenetic case reports, papers on descriptive population genetics, articles dealing with the frequency of polymorphisms or additional mutations within genes in which numerous lesions have already been described, and papers that report meta-analyses of previously published datasets will normally not be accepted.
The Journal typically will not consider for publication manuscripts that report merely the isolation, map position, structure, and tissue expression profile of a gene of unknown function unless the gene is of particular interest or is a candidate gene involved in a human trait or disorder.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


