{"title":"Targeting iron–sulfur cluster assembly scaffold protein as a vulnerability in KRAS-activated pancreatic ductal adenocarcinoma","authors":"Huarui Cai, Hongjuan Cui, Erhu Zhao","doi":"10.1002/mog2.65","DOIUrl":null,"url":null,"abstract":"<p>In two recent publications in Science<span><sup>1</sup></span> and Cell Discovery,<span><sup>2</sup></span> researchers have discovered that iron–sulfur cluster assembly scaffold protein (ISCU) plays crucial roles on maintaining glutathione (GSH) homeostasis, α-KG catabolism, DNA methylation and so on. These groundbreaking findings not only establish crucial connections among iron–sulfur (Fe–S) metabolism, GSH homeostasis, alpha-ketoglutarate (α-KG) catabolism, DNA methylation, and tumor growth but also emphasize the significant and integrated regulation of mitochondrial function and gene expression by ISCU (Figure 1).</p><p>Liu et al.<span><sup>1</sup></span> unveiled the vital role of ISCU in maintaining optimal mitochondrial functions by restoring the balance between iron and GSH. In addition, a previous study<span><sup>3</sup></span> has provided evidence that glutamine, a precursor of GSH, promotes tumor growth through a KRAS-regulated metabolic pathway in pancreatic ductal adenocarcinoma (PDAC), thereby suggesting that ISCU potentially influences tumor development by modulating GSH homeostasis. Ren et al.<span><sup>2</sup></span> uncovered a previously undisclosed association between Fe–S metabolism and tumor growth in kirsten rat sarcoma viral oncogene homolog (KRAS)-activated PDAC. Such findings offer a promising avenue for the treatment of KRAS-activated PDAC, as well as for interventions targeting ISCU-mediated cellular processes.</p><p>PDAC is a highly lethal disease with a mounting incidence and unfavorable prognostic outcomes, underscoring the urgent requirement for the development of efficacious therapeutic approaches. Thus, accurate biomarkers to help stratify risk would greatly improve current diagnostic and decision-making dilemmas. Fe–S clusters are ancient, ubiquitous metal cofactors that possess various physiological functions in antioxidant, iron regulation, the tricarboxylic acid (TCA) cycle, and many other metabolic reactions. Accumulated evidence suggests that abnormal Fe–S clusters assembly pathways lead to mitochondrial dysfunction and cause various diseases, particularly with an impact on tumor development.<span><sup>4</sup></span> For instance, nitrogen fixation gene 1 (NFS1) cysteine desulfurase, as an indispensable protein in Fe–S cluster biogenesis, plays a vital role in mitochondrial metabolic reprogramming. In human colorectal cancer, NFS1 can suppress PANoptosis under oxaliplatin chemotherapy, and NFS1 high expression was linked to unfavorable survival outcomes and hyposensitivity to chemotherapy in patients.<span><sup>5</sup></span> In addition, succinate dehydrogenase complex iron–sulfur subunit B (SDHB) is also an Fe–S cluster protein consisting of three Fe–S clusters. In SDHB-deficient cancer cells, succinate levels are elevated, resulting in hypermethylation of histones and DNA, and glutamine becomes the primary source of TCA cycle metabolites via reductive carboxylation.<span><sup>6</sup></span></p><p>Notably, ISCU, a key component of the core Fe–S cluster assembly (ISC) complexes, is instrumental in synthesizing a bridging [2Fe–2S] cluster on dimeric ISCU in human mitochondria. Liu et al.<span><sup>1</sup></span> have reported that ISCU acts as a regulator of the mitochondrial GSH transporter, solute carrier family 25 member 39 (SLC25A39), which is essential for maintaining redox balance and ensuring the proper equilibrium between iron availability and GSH levels within the mitochondria. And KRAS-mediated GSH homeostasis have been found to be critical for maintaining redox balance and promoting cell growth in PDAC.<span><sup>3</sup></span> Furthermore, Ren et al.<span><sup>2</sup></span> reveals that ISCU, activated by KRAS-enhanced MYC expression in PDAC cells, promotes α-KG catabolism and mitochondrial functions accompanied by DNA 5-methylcytosine (5mC)-dependent cell proliferation and tumor growth. Collectively, ISCU is expected to be a potential biomarker for diagnosis, surveillance, and therapy of PDAC.</p><p>Alpha-KG serves as a critical intermediate metabolite in the TCA cycle, linking multiple metabolic pathways and various cellular functions in cancer. Ren et al.<span><sup>2</sup></span> have revealed that KRAS can significantly regulate the absolute content of α-KG in cells through in vitro and in vivo experiments, and changes in α-KG levels caused by KRAS silencing or overexpression of KRAS G12V can be reversed by depleting glutamine in the culture medium. Moreover, it was further confirmed that KRAS enhances α-KG catabolism, rather than reducing α-KG production from glutamate, which serves as a fundamental component for GSH synthesis.<span><sup>7</sup></span> Additionally, Liu et al.<span><sup>1</sup></span> discovered that GSH, a crucial endogenous ligand for iron, is involved in a process where depletion of GSH results in a strong binding of a [2Fe–2S] cluster to SLC25A39 via cysteines (C74/78/88/94). ISCU loss has been demonstrated to abolish the stabilization of SLC25A39 under GSH depletion. Conversely, loss of ATP binding cassette subfamily B member 7, a putative transporter involved in exporting Fe–S clusters from mitochondria, has been found to consistently stabilize SLC25A39. This stabilization is likely due to the accumulation of [2Fe–2S] clusters within the mitochondria. Finally, ISCU has been shown to play a crucial role in maintaining optimal mitochondrial functions by restoring iron/GSH balance through the adaptive response to iron and GSH equilibrium of SLC25A39.</p><p>Moreover, Ren et al.<span><sup>2</sup></span> further confirmed the deletion of ISCU suppressed the expression of all Fe–S dependent mitochondrial proteins, including dihydrolipoamide <i>S</i>-succinyltransferase (DLST) and aconitase 2 (ACO2) in PDAC cells. Notably, KRAS solely changed the protein level of DLST that is stable with lipoylation, without its messenger RNA level. Subsequent investigations showed that the decrease in the DLST protein levels and ACO2 activity caused by KRAS loss could be counteracted by ISCU overexpression. Although the decrease of ISCU caused by KRAS silencing was not substantial enough to suppress ACO2 protein levels, it was sufficient to cause a decrease in ACO2 activity, suggesting that the stability of ACO2, controlled by ISCU, only diminishes under a specific threshold of Fe–S clusters.</p><p>As a primary metabolite, α-KG serves various functions, such as a vital cofactor for the dioxygenase enzymes: 10–11 translocation methylcytosine dioxygenase 1,2,3 (TET1, TET2, TET3), which can catalyze 5mC oxidation and regulate DNA methylation. Ren et al.<span><sup>2</sup></span> revealed that activated KRAS enhances the DNA methylation by the control of ISCU-promoted α-KG catabolism and TET3 inhibition. The alterations in DNA 5mC methylation are associated with the development of diseases, especially malignant tumor.<span><sup>8</sup></span> Therefore, the increase of ISCU-mediated DNA 5mC level may alter the expression of numerous genes. Ren et al.<span><sup>2</sup></span> found that 103 differentially expressed genes altered by ISCU silencing through performing the whole-genome bisulfite sequencing. Among them, it is worth noting that DNA polymerase I (POLA1) possesses the capability to initiate DNA synthesis during the S phase of the cell cycle. Finally, they confirmed that dysregulation of 5mC methylation caused an aberrant transcription (as exemplified by the activation of POLA1), which may contribute to the oncogenic transformation of PDAC.</p><p>Moreover, by adding α-KG competitive metabolic inhibitors and creating hypoxic conditions to mitigate electron transport chain dysregulation, Ren et al.<span><sup>2</sup></span> found that ISCU controlled cell proliferation in an α-KG-dependent manner. Certain clinical drugs such as decitabine, used to treat myelodysplastic syndrome are also DNA methyltransferase (DNMT) inhibitors.<span><sup>9</sup></span> Ren et al.<span><sup>2</sup></span> found that ISCU low-expression cells were insensitive to decitabine, suggesting that the expression level of ISCU has the potential to serve as a molecular biomarker for the treatment of PDAC with DNMT inhibitors.</p><p>In summary, Liu et al.<span><sup>1</sup></span> and Ren et al.<span><sup>2</sup></span> have provided compelling evidence for the crucial roles of ISCU in maintaining GSH homeostasis, α-KG catabolism and DNA methylation, and so on. Moreover, the authors have uncovered an intriguing connection between Fe–S metabolism and tumor growth in PDAC, offering new insights into the pathogenesis of PDAC, particularly in KRAS-activated mutant cancer subtypes. Remarkably, ISCU has been identified as a regulator of DNA 5mC level in KRAS-activated PDAC. These findings not only propose the potential repurposing of existing DNMT inhibitors but also open doors for the development of novel inhibitors, presenting a promising avenue for clinical intervention in PDAC.</p><p><b>Huarui Cai:</b> Conceptualization (equal); visualization (lead); writing—original draft (lead). <b>Hongjuan Cui:</b> Supervision (equal); writing—review and editing (supporting). <b>Erhu Zhao:</b> Conceptualization (equal); funding acquisition (lead); supervision (equal); writing—review and editing (lead). The final manuscript has been approved by all authors.</p><p>The authors declare no conflicts of interest.</p><p>Not applicable.</p>","PeriodicalId":100902,"journal":{"name":"MedComm – Oncology","volume":"3 1","pages":""},"PeriodicalIF":0.0000,"publicationDate":"2024-01-29","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/mog2.65","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"MedComm – Oncology","FirstCategoryId":"1085","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/mog2.65","RegionNum":0,"RegionCategory":null,"ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"","JCRName":"","Score":null,"Total":0}
引用次数: 0
Abstract
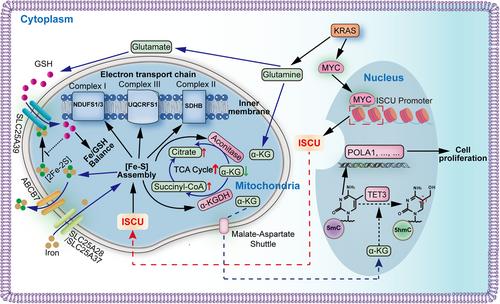
In two recent publications in Science1 and Cell Discovery,2 researchers have discovered that iron–sulfur cluster assembly scaffold protein (ISCU) plays crucial roles on maintaining glutathione (GSH) homeostasis, α-KG catabolism, DNA methylation and so on. These groundbreaking findings not only establish crucial connections among iron–sulfur (Fe–S) metabolism, GSH homeostasis, alpha-ketoglutarate (α-KG) catabolism, DNA methylation, and tumor growth but also emphasize the significant and integrated regulation of mitochondrial function and gene expression by ISCU (Figure 1).
Liu et al.1 unveiled the vital role of ISCU in maintaining optimal mitochondrial functions by restoring the balance between iron and GSH. In addition, a previous study3 has provided evidence that glutamine, a precursor of GSH, promotes tumor growth through a KRAS-regulated metabolic pathway in pancreatic ductal adenocarcinoma (PDAC), thereby suggesting that ISCU potentially influences tumor development by modulating GSH homeostasis. Ren et al.2 uncovered a previously undisclosed association between Fe–S metabolism and tumor growth in kirsten rat sarcoma viral oncogene homolog (KRAS)-activated PDAC. Such findings offer a promising avenue for the treatment of KRAS-activated PDAC, as well as for interventions targeting ISCU-mediated cellular processes.
PDAC is a highly lethal disease with a mounting incidence and unfavorable prognostic outcomes, underscoring the urgent requirement for the development of efficacious therapeutic approaches. Thus, accurate biomarkers to help stratify risk would greatly improve current diagnostic and decision-making dilemmas. Fe–S clusters are ancient, ubiquitous metal cofactors that possess various physiological functions in antioxidant, iron regulation, the tricarboxylic acid (TCA) cycle, and many other metabolic reactions. Accumulated evidence suggests that abnormal Fe–S clusters assembly pathways lead to mitochondrial dysfunction and cause various diseases, particularly with an impact on tumor development.4 For instance, nitrogen fixation gene 1 (NFS1) cysteine desulfurase, as an indispensable protein in Fe–S cluster biogenesis, plays a vital role in mitochondrial metabolic reprogramming. In human colorectal cancer, NFS1 can suppress PANoptosis under oxaliplatin chemotherapy, and NFS1 high expression was linked to unfavorable survival outcomes and hyposensitivity to chemotherapy in patients.5 In addition, succinate dehydrogenase complex iron–sulfur subunit B (SDHB) is also an Fe–S cluster protein consisting of three Fe–S clusters. In SDHB-deficient cancer cells, succinate levels are elevated, resulting in hypermethylation of histones and DNA, and glutamine becomes the primary source of TCA cycle metabolites via reductive carboxylation.6
Notably, ISCU, a key component of the core Fe–S cluster assembly (ISC) complexes, is instrumental in synthesizing a bridging [2Fe–2S] cluster on dimeric ISCU in human mitochondria. Liu et al.1 have reported that ISCU acts as a regulator of the mitochondrial GSH transporter, solute carrier family 25 member 39 (SLC25A39), which is essential for maintaining redox balance and ensuring the proper equilibrium between iron availability and GSH levels within the mitochondria. And KRAS-mediated GSH homeostasis have been found to be critical for maintaining redox balance and promoting cell growth in PDAC.3 Furthermore, Ren et al.2 reveals that ISCU, activated by KRAS-enhanced MYC expression in PDAC cells, promotes α-KG catabolism and mitochondrial functions accompanied by DNA 5-methylcytosine (5mC)-dependent cell proliferation and tumor growth. Collectively, ISCU is expected to be a potential biomarker for diagnosis, surveillance, and therapy of PDAC.
Alpha-KG serves as a critical intermediate metabolite in the TCA cycle, linking multiple metabolic pathways and various cellular functions in cancer. Ren et al.2 have revealed that KRAS can significantly regulate the absolute content of α-KG in cells through in vitro and in vivo experiments, and changes in α-KG levels caused by KRAS silencing or overexpression of KRAS G12V can be reversed by depleting glutamine in the culture medium. Moreover, it was further confirmed that KRAS enhances α-KG catabolism, rather than reducing α-KG production from glutamate, which serves as a fundamental component for GSH synthesis.7 Additionally, Liu et al.1 discovered that GSH, a crucial endogenous ligand for iron, is involved in a process where depletion of GSH results in a strong binding of a [2Fe–2S] cluster to SLC25A39 via cysteines (C74/78/88/94). ISCU loss has been demonstrated to abolish the stabilization of SLC25A39 under GSH depletion. Conversely, loss of ATP binding cassette subfamily B member 7, a putative transporter involved in exporting Fe–S clusters from mitochondria, has been found to consistently stabilize SLC25A39. This stabilization is likely due to the accumulation of [2Fe–2S] clusters within the mitochondria. Finally, ISCU has been shown to play a crucial role in maintaining optimal mitochondrial functions by restoring iron/GSH balance through the adaptive response to iron and GSH equilibrium of SLC25A39.
Moreover, Ren et al.2 further confirmed the deletion of ISCU suppressed the expression of all Fe–S dependent mitochondrial proteins, including dihydrolipoamide S-succinyltransferase (DLST) and aconitase 2 (ACO2) in PDAC cells. Notably, KRAS solely changed the protein level of DLST that is stable with lipoylation, without its messenger RNA level. Subsequent investigations showed that the decrease in the DLST protein levels and ACO2 activity caused by KRAS loss could be counteracted by ISCU overexpression. Although the decrease of ISCU caused by KRAS silencing was not substantial enough to suppress ACO2 protein levels, it was sufficient to cause a decrease in ACO2 activity, suggesting that the stability of ACO2, controlled by ISCU, only diminishes under a specific threshold of Fe–S clusters.
As a primary metabolite, α-KG serves various functions, such as a vital cofactor for the dioxygenase enzymes: 10–11 translocation methylcytosine dioxygenase 1,2,3 (TET1, TET2, TET3), which can catalyze 5mC oxidation and regulate DNA methylation. Ren et al.2 revealed that activated KRAS enhances the DNA methylation by the control of ISCU-promoted α-KG catabolism and TET3 inhibition. The alterations in DNA 5mC methylation are associated with the development of diseases, especially malignant tumor.8 Therefore, the increase of ISCU-mediated DNA 5mC level may alter the expression of numerous genes. Ren et al.2 found that 103 differentially expressed genes altered by ISCU silencing through performing the whole-genome bisulfite sequencing. Among them, it is worth noting that DNA polymerase I (POLA1) possesses the capability to initiate DNA synthesis during the S phase of the cell cycle. Finally, they confirmed that dysregulation of 5mC methylation caused an aberrant transcription (as exemplified by the activation of POLA1), which may contribute to the oncogenic transformation of PDAC.
Moreover, by adding α-KG competitive metabolic inhibitors and creating hypoxic conditions to mitigate electron transport chain dysregulation, Ren et al.2 found that ISCU controlled cell proliferation in an α-KG-dependent manner. Certain clinical drugs such as decitabine, used to treat myelodysplastic syndrome are also DNA methyltransferase (DNMT) inhibitors.9 Ren et al.2 found that ISCU low-expression cells were insensitive to decitabine, suggesting that the expression level of ISCU has the potential to serve as a molecular biomarker for the treatment of PDAC with DNMT inhibitors.
In summary, Liu et al.1 and Ren et al.2 have provided compelling evidence for the crucial roles of ISCU in maintaining GSH homeostasis, α-KG catabolism and DNA methylation, and so on. Moreover, the authors have uncovered an intriguing connection between Fe–S metabolism and tumor growth in PDAC, offering new insights into the pathogenesis of PDAC, particularly in KRAS-activated mutant cancer subtypes. Remarkably, ISCU has been identified as a regulator of DNA 5mC level in KRAS-activated PDAC. These findings not only propose the potential repurposing of existing DNMT inhibitors but also open doors for the development of novel inhibitors, presenting a promising avenue for clinical intervention in PDAC.
Huarui Cai: Conceptualization (equal); visualization (lead); writing—original draft (lead). Hongjuan Cui: Supervision (equal); writing—review and editing (supporting). Erhu Zhao: Conceptualization (equal); funding acquisition (lead); supervision (equal); writing—review and editing (lead). The final manuscript has been approved by all authors.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


