Henry S. Pollock, Corey E. Tarwater, James R. Karr, Jeffrey D. Brawn
{"title":"长期监测显示,新热带森林陆鸟的寿命很长。","authors":"Henry S. Pollock, Corey E. Tarwater, James R. Karr, Jeffrey D. Brawn","doi":"10.1002/ecy.4386","DOIUrl":null,"url":null,"abstract":"<p>Longevity, or maximum lifespan, is a trait that has fascinated ecologists, evolutionary biologists, and geneticists for centuries and has featured prominently in the development of life history, aging, and senescence theories (Lankester, <span>1870</span>; Nussey et al., <span>2013</span>; Stearns, <span>1992</span>). Longevity has been associated with a variety of other life history and morphological traits such as brain and body size (Lindstedt & Calder, <span>1981</span>; Stearns, <span>1992</span>), energy metabolism (Furness & Speakman, <span>2008</span>; White & Seymour, <span>2004</span>), hibernation strategy (Turbill et al., <span>2011</span>; Wilkinson & South, <span>2002</span>), and antioxidant capacity (Vágási et al., <span>2019</span>). Meaningful longevity data, however, necessitate studies of marked individuals over extended periods, a requirement that can be challenging for long-lived vertebrate species. Long-term mark-recapture efforts provide some of the best opportunities for obtaining longevity estimates from free-living populations of animals. For example, among birds, continent-wide banding programs in North America (i.e., U.S.G.S. Bird Banding Laboratory [BBL]: https://www.pwrc.usgs.gov/bbl/longevity/longevity_main.cfm) and Europe (i.e., EURING: https://euring.org/data-and-codes/longevity-list) comprise decades of individual-based monitoring that have yielded longevity estimates for many temperate-zone species. By contrast, longevity data from tropical birds are few and restricted in temporal scope due to the logistical and financial challenges of maintaining long-term monitoring programs in the tropics (Collen et al., <span>2008</span>; Proença et al., <span>2017</span>). As of 2023, fewer than 250 longevity records of Neotropical landbirds are available in the published literature (reviewed in Scholer et al., <span>2018</span>), and only two studies spanned ≥20 years (Arendt, <span>2006</span>; Woodworth et al., <span>1999</span>).</p><p>How long do Neotropical forest landbirds actually live? To address this question, we used a 45-year (1977–2022) mark-recapture dataset from central Panama—one of the longest of its kind in the Neotropics—to derive longevity records for 47 species. We sampled birds on the 100-ha Limbo Plot on Pipeline Road in the 22,000-ha Soberanía National Park (9.15° N, 79.73° W). The Limbo Plot is classified as tropical lowland moist forest and has been the focus of numerous previous studies of tropical bird ecology (e.g., Brawn et al., <span>2017</span>; Karr, <span>1976</span>; Karr & Freemark, <span>1983</span>; Pollock et al., <span>2020</span>, <span>2022</span>; Tarwater et al., <span>2011</span>; see Robinson et al., <span>2000</span> for a more detailed discussion of the Limbo Plot). Netting and banding protocols are described in detail in Pollock et al. (<span>2022</span>). Briefly, we mist-netted and banded birds twice per year (dry and wet season) at two 2-ha sites embedded in the larger 100-ha Limbo Plot. At each netting site, nets were set at ground level and kept open for approximately 600 total net-hours each sampling period. To estimate longevity, we compiled capture histories of more than 15,000 individuals of 150 species, indexed by unique band numbers. We excluded species with ≤20 total captures over the 45-year study period, as they were unlikely to use the understory stratum sampled by mist-nets or too locally rare to yield accurate longevity estimates. We then derived longevity records for 198 individuals of 47 tropical bird species in 17 families (henRyPollock, <span>2024</span> at https://doi.org/10.5281/zenodo.11106481). We report the minimum longevity (i.e., the elapsed time between initial banding and last recapture, <i>sensu</i> Scholer et al., <span>2018</span>) of the longest-lived individual for each of the 47 species (Appendix S1: Table S1).</p><p>The oldest individual longevity record was an 18-year-old Rufous Motmot (<i>Baryphthengus martii</i>; Figure 1A). Other notably long-lived individuals included a Plain-brown Woodcreeper (<i>Dendrocincla fuliginosa</i>; 15.7 years; Figure 1B), two Golden-crowned Spadebills (<i>Platyrinchus coronatus</i>; 14.0 and 14.3 years; Figure 1C), two Bright-rumped Attilas (<i>Attila spadiceus</i>; 12.0 and 14.2 years; Figure 1D), an Ocellated Antbird (<i>Phaenostictus mcleannani</i>; 12.4 years; Figure 1E), and a Black-throated Trogon (<i>Trogon rufus</i>; 12.0 years; Figure 1F). Of the 47 longevity estimates included in the dataset (Figure 2A), 37 (79%) were the longest on record and 27 (57%) were first records for a given species, respectively (Appendix S1: Table S1). Most species (34/47; 72%) had a longevity of ≥7 years, 29 (62%) had a longevity of ≥8 years, and 18 (38%) had a longevity of ≥10 years (Appendix S1: Figure S1).</p><p>Of the 17 families included in the dataset, 14 (82%) had at least one species with a longevity record of ≥8 years and 10 (59%) had at least one species with a longevity record of ≥10 years (Figure 2B). Families with high longevities included the ovenbirds (Furnariidae: 6 of 9 species with ≥8 years longevity) and the antbirds (Thamnophilidae: all nine species ≥7 years; 7 of 9 species with ≥8 years longevity). Flycatchers (Tyrannidae) had a bimodal distribution, with very high longevities for some species (e.g., >14 years; Bright-rumped Attila and Golden-crowned Spadebill) and relatively low longevities (<7 years) for most other species despite similar recapture rates (Appendix S1: Table S1; henRyPollock, <span>2024</span>). We tested for and found no significant differences in longevity between oscine passerines, suboscine passerines, and non-passerines (generalized linear model [GLM]: <i>F</i><sub>2,44</sub> = 2.0, <i>p</i> = 0.15, <i>R</i><sup>2</sup> = 0.08; Appendix S1: Figure S2).</p><p>We also tested for potential relationships between species' longevities and ecological traits including body mass, diet (i.e., frugivore, insectivore, omnivore, carnivore), and sociality (i.e., non-flocker, facultative flocker, obligate flocker, ant-follower, lekking species). We found no significant associations between longevity and body mass (GLM: <i>F</i><sub>1,45</sub> = 0.84, <i>p</i> = 0.36, <i>R</i><sup>2</sup> = 0.02; Appendix S1: Figure S3), diet (GLM: <i>F</i><sub>4,42</sub> = 1.0, <i>p</i> = 0.42, <i>R</i><sup>2</sup> = 0.09), or sociality (GLM: <i>F</i><sub>4,42</sub> = 1.10, <i>p</i> = 0.37, <i>R</i><sup>2</sup> = 0.09). Individuals captured more frequently or with higher abundances were expected to have higher longevity estimates, but we found no relationship between longevity and the number of times an individual was captured (GLM: <i>F</i><sub>1,196</sub> = 2.0, <i>p</i> = 0.16, <i>R</i><sup>2</sup> = 0.01; Appendix S1: Figure S4) or the total number of captures per species (GLM: <i>F</i><sub>1,39</sub> = 0.82, <i>p</i> = 0.37, <i>R</i><sup>2</sup> = 0.02). Finally, longevity has been questioned for its value as a life history trait versus, for example, annual survival rate (Krementz et al., <span>1989</span>). Therefore, we tested for and found a moderately high positive correlation for 18 species from our study site for which estimates of both annual survival rates (derived from Brawn et al., <span>2017</span>) and longevities were available (GLM: <i>F</i><sub>1,16</sub> = 3.05, <i>p</i> < 0.001, <i>R</i><sup>2</sup> = 0.50; Appendix S1: Figure S5).</p><p>The duration and depth of our study (45 years, >80,000 captures) provided an unprecedented window into the lives of Neotropical forest birds and revealed that many Neotropical species have considerably longer lifespans than previously documented in the scientific literature. We report the oldest longevity records to date for ~80% of all species and ~65% of all families sampled, encompassing a broad range of phylogenetic and ecological variation. Surprisingly, and in contrast with other studies (e.g., Furness & Speakman, <span>2008</span>; Lindstedt & Calder, <span>1981</span>), we found no correlation between body mass and longevity, perhaps because our focal species comprised a relatively narrow range of body masses (7–219 g; Appendix S1: Table S1) relative to other datasets. In fact, some of the smallest-bodied species (<15 g) at our study site were also the longest-lived, including Plain Xenops (<i>Xenops minutus</i>—11.0 years), White-flanked Antwren (<i>Myrmotherula axillaris</i>—11.0 years) and Golden-crowned Spadebill (<i>Platyrinchus coronatus</i>—14.3 years). Finally, non-passerines are thought to have longer lifespans (Lindstedt & Calder, <span>1976</span>), but we found no evidence of differences in longevity between passerines and non-passerines, which we expect was due to having few non-passerines (<i>n</i> = 6 species) in our dataset, all of which had relatively small body sizes (42–219 g). Overall, our results reveal the long lifespans of tropical landbirds and reinforce the importance of long-term studies for obtaining these types of data (Magurran et al., <span>2010</span>).</p><p>Nevertheless, several lines of evidence suggest that our longevity records are still likely biased underestimates of true longevity. First, the limited spatial extent of our study (two 2-ha plots) means that many individuals that were not recaptured could still have been persisting undetected outside of the study plots. Second, longevity was not correlated with the average number of recaptures of the sampled species, and nearly 60% of longevity records were based on individuals recaptured five or fewer times. For example, species with large home-ranges like army-ant-following birds (e.g., Ocellated Antbird—<i>Phaenostictus mcleannani</i>) and non-understory species (e.g., Yellow-margined Flycatcher—<i>Tolmomyias assimilis</i>) were captured less frequently, and continued sampling will likely yield longer longevity records for these species in particular. Finally, 11 of our longevity records were lower than those reported in other studies of shorter duration (Appendix S1: Table S1). In particular, studies of color-banded populations that have tracked individuals over long time periods (e.g., Tarwater & Kelley, <span>2020</span>; Willis, <span>1983</span>) have yielded longer longevity records than our community-level mark-recapture study, likely due to greater sampling effort and higher detection probability of color-banded individuals. Thus, our data suggest that even 40+ years of mark-recapture effort may not be sufficient to produce accurate longevity estimates for certain Neotropical forest bird species.</p><p>Future work that endeavors to address questions about how longevity varies geographically and assess its ecological relevance as a life history trait may require large-scale collaborative banding schemes and long-term population studies of banded birds in tropical regions to obtain broader geographical coverage and more accurate longevity estimates. Currently, the differences in spatial coverage and number of captures between our dataset (one site, thousands of captures) and large-scale banding schemes such as the BBL in North America and EURING in Europe (thousands of sites, millions of captures) span orders of magnitude, making direct comparison challenging. One potential way to test for geographical differences in longevity is standardized comparisons from single-site banding operations (e.g., banding sites included in the Institute for Bird Populations' Monitoring Avian Productivity and Survivorship: https://www.birdpop.org/pages/maps.php) across similar time periods. These long-term datasets are particularly difficult to compile in tropics, however, where biodiversity is greatest but sampling effort still lags behind that of temperate systems (Collen et al., <span>2008</span>). For example, Scholer et al. (<span>2018</span>) found that avian longevity estimates of Neotropical landbirds were highly correlated with study duration, highlighting the need for longer-term studies and the systemic challenges to amassing such longevity datasets in the tropics. Therefore, despite the moderately high correlation (<i>R</i><sup>2</sup> = 0.5) we found between survival and longevity, we concur with the conclusions of Krementz et al. (<span>1989</span>), who found that model-based estimates of survival were preferable to observed maximum lifespan statistics for use in comparative life history studies.</p><p>The authors declare no conflicts of interest.</p>","PeriodicalId":11484,"journal":{"name":"Ecology","volume":"105 9","pages":""},"PeriodicalIF":4.4000,"publicationDate":"2024-07-24","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ecy.4386","citationCount":"0","resultStr":"{\"title\":\"Long-term monitoring reveals the long lifespans of Neotropical forest landbirds\",\"authors\":\"Henry S. Pollock, Corey E. Tarwater, James R. Karr, Jeffrey D. Brawn\",\"doi\":\"10.1002/ecy.4386\",\"DOIUrl\":null,\"url\":null,\"abstract\":\"<p>Longevity, or maximum lifespan, is a trait that has fascinated ecologists, evolutionary biologists, and geneticists for centuries and has featured prominently in the development of life history, aging, and senescence theories (Lankester, <span>1870</span>; Nussey et al., <span>2013</span>; Stearns, <span>1992</span>). Longevity has been associated with a variety of other life history and morphological traits such as brain and body size (Lindstedt & Calder, <span>1981</span>; Stearns, <span>1992</span>), energy metabolism (Furness & Speakman, <span>2008</span>; White & Seymour, <span>2004</span>), hibernation strategy (Turbill et al., <span>2011</span>; Wilkinson & South, <span>2002</span>), and antioxidant capacity (Vágási et al., <span>2019</span>). Meaningful longevity data, however, necessitate studies of marked individuals over extended periods, a requirement that can be challenging for long-lived vertebrate species. Long-term mark-recapture efforts provide some of the best opportunities for obtaining longevity estimates from free-living populations of animals. For example, among birds, continent-wide banding programs in North America (i.e., U.S.G.S. Bird Banding Laboratory [BBL]: https://www.pwrc.usgs.gov/bbl/longevity/longevity_main.cfm) and Europe (i.e., EURING: https://euring.org/data-and-codes/longevity-list) comprise decades of individual-based monitoring that have yielded longevity estimates for many temperate-zone species. By contrast, longevity data from tropical birds are few and restricted in temporal scope due to the logistical and financial challenges of maintaining long-term monitoring programs in the tropics (Collen et al., <span>2008</span>; Proença et al., <span>2017</span>). As of 2023, fewer than 250 longevity records of Neotropical landbirds are available in the published literature (reviewed in Scholer et al., <span>2018</span>), and only two studies spanned ≥20 years (Arendt, <span>2006</span>; Woodworth et al., <span>1999</span>).</p><p>How long do Neotropical forest landbirds actually live? To address this question, we used a 45-year (1977–2022) mark-recapture dataset from central Panama—one of the longest of its kind in the Neotropics—to derive longevity records for 47 species. We sampled birds on the 100-ha Limbo Plot on Pipeline Road in the 22,000-ha Soberanía National Park (9.15° N, 79.73° W). The Limbo Plot is classified as tropical lowland moist forest and has been the focus of numerous previous studies of tropical bird ecology (e.g., Brawn et al., <span>2017</span>; Karr, <span>1976</span>; Karr & Freemark, <span>1983</span>; Pollock et al., <span>2020</span>, <span>2022</span>; Tarwater et al., <span>2011</span>; see Robinson et al., <span>2000</span> for a more detailed discussion of the Limbo Plot). Netting and banding protocols are described in detail in Pollock et al. (<span>2022</span>). Briefly, we mist-netted and banded birds twice per year (dry and wet season) at two 2-ha sites embedded in the larger 100-ha Limbo Plot. At each netting site, nets were set at ground level and kept open for approximately 600 total net-hours each sampling period. To estimate longevity, we compiled capture histories of more than 15,000 individuals of 150 species, indexed by unique band numbers. We excluded species with ≤20 total captures over the 45-year study period, as they were unlikely to use the understory stratum sampled by mist-nets or too locally rare to yield accurate longevity estimates. We then derived longevity records for 198 individuals of 47 tropical bird species in 17 families (henRyPollock, <span>2024</span> at https://doi.org/10.5281/zenodo.11106481). We report the minimum longevity (i.e., the elapsed time between initial banding and last recapture, <i>sensu</i> Scholer et al., <span>2018</span>) of the longest-lived individual for each of the 47 species (Appendix S1: Table S1).</p><p>The oldest individual longevity record was an 18-year-old Rufous Motmot (<i>Baryphthengus martii</i>; Figure 1A). Other notably long-lived individuals included a Plain-brown Woodcreeper (<i>Dendrocincla fuliginosa</i>; 15.7 years; Figure 1B), two Golden-crowned Spadebills (<i>Platyrinchus coronatus</i>; 14.0 and 14.3 years; Figure 1C), two Bright-rumped Attilas (<i>Attila spadiceus</i>; 12.0 and 14.2 years; Figure 1D), an Ocellated Antbird (<i>Phaenostictus mcleannani</i>; 12.4 years; Figure 1E), and a Black-throated Trogon (<i>Trogon rufus</i>; 12.0 years; Figure 1F). Of the 47 longevity estimates included in the dataset (Figure 2A), 37 (79%) were the longest on record and 27 (57%) were first records for a given species, respectively (Appendix S1: Table S1). Most species (34/47; 72%) had a longevity of ≥7 years, 29 (62%) had a longevity of ≥8 years, and 18 (38%) had a longevity of ≥10 years (Appendix S1: Figure S1).</p><p>Of the 17 families included in the dataset, 14 (82%) had at least one species with a longevity record of ≥8 years and 10 (59%) had at least one species with a longevity record of ≥10 years (Figure 2B). Families with high longevities included the ovenbirds (Furnariidae: 6 of 9 species with ≥8 years longevity) and the antbirds (Thamnophilidae: all nine species ≥7 years; 7 of 9 species with ≥8 years longevity). Flycatchers (Tyrannidae) had a bimodal distribution, with very high longevities for some species (e.g., >14 years; Bright-rumped Attila and Golden-crowned Spadebill) and relatively low longevities (<7 years) for most other species despite similar recapture rates (Appendix S1: Table S1; henRyPollock, <span>2024</span>). We tested for and found no significant differences in longevity between oscine passerines, suboscine passerines, and non-passerines (generalized linear model [GLM]: <i>F</i><sub>2,44</sub> = 2.0, <i>p</i> = 0.15, <i>R</i><sup>2</sup> = 0.08; Appendix S1: Figure S2).</p><p>We also tested for potential relationships between species' longevities and ecological traits including body mass, diet (i.e., frugivore, insectivore, omnivore, carnivore), and sociality (i.e., non-flocker, facultative flocker, obligate flocker, ant-follower, lekking species). We found no significant associations between longevity and body mass (GLM: <i>F</i><sub>1,45</sub> = 0.84, <i>p</i> = 0.36, <i>R</i><sup>2</sup> = 0.02; Appendix S1: Figure S3), diet (GLM: <i>F</i><sub>4,42</sub> = 1.0, <i>p</i> = 0.42, <i>R</i><sup>2</sup> = 0.09), or sociality (GLM: <i>F</i><sub>4,42</sub> = 1.10, <i>p</i> = 0.37, <i>R</i><sup>2</sup> = 0.09). Individuals captured more frequently or with higher abundances were expected to have higher longevity estimates, but we found no relationship between longevity and the number of times an individual was captured (GLM: <i>F</i><sub>1,196</sub> = 2.0, <i>p</i> = 0.16, <i>R</i><sup>2</sup> = 0.01; Appendix S1: Figure S4) or the total number of captures per species (GLM: <i>F</i><sub>1,39</sub> = 0.82, <i>p</i> = 0.37, <i>R</i><sup>2</sup> = 0.02). Finally, longevity has been questioned for its value as a life history trait versus, for example, annual survival rate (Krementz et al., <span>1989</span>). Therefore, we tested for and found a moderately high positive correlation for 18 species from our study site for which estimates of both annual survival rates (derived from Brawn et al., <span>2017</span>) and longevities were available (GLM: <i>F</i><sub>1,16</sub> = 3.05, <i>p</i> < 0.001, <i>R</i><sup>2</sup> = 0.50; Appendix S1: Figure S5).</p><p>The duration and depth of our study (45 years, >80,000 captures) provided an unprecedented window into the lives of Neotropical forest birds and revealed that many Neotropical species have considerably longer lifespans than previously documented in the scientific literature. We report the oldest longevity records to date for ~80% of all species and ~65% of all families sampled, encompassing a broad range of phylogenetic and ecological variation. Surprisingly, and in contrast with other studies (e.g., Furness & Speakman, <span>2008</span>; Lindstedt & Calder, <span>1981</span>), we found no correlation between body mass and longevity, perhaps because our focal species comprised a relatively narrow range of body masses (7–219 g; Appendix S1: Table S1) relative to other datasets. In fact, some of the smallest-bodied species (<15 g) at our study site were also the longest-lived, including Plain Xenops (<i>Xenops minutus</i>—11.0 years), White-flanked Antwren (<i>Myrmotherula axillaris</i>—11.0 years) and Golden-crowned Spadebill (<i>Platyrinchus coronatus</i>—14.3 years). Finally, non-passerines are thought to have longer lifespans (Lindstedt & Calder, <span>1976</span>), but we found no evidence of differences in longevity between passerines and non-passerines, which we expect was due to having few non-passerines (<i>n</i> = 6 species) in our dataset, all of which had relatively small body sizes (42–219 g). Overall, our results reveal the long lifespans of tropical landbirds and reinforce the importance of long-term studies for obtaining these types of data (Magurran et al., <span>2010</span>).</p><p>Nevertheless, several lines of evidence suggest that our longevity records are still likely biased underestimates of true longevity. First, the limited spatial extent of our study (two 2-ha plots) means that many individuals that were not recaptured could still have been persisting undetected outside of the study plots. Second, longevity was not correlated with the average number of recaptures of the sampled species, and nearly 60% of longevity records were based on individuals recaptured five or fewer times. For example, species with large home-ranges like army-ant-following birds (e.g., Ocellated Antbird—<i>Phaenostictus mcleannani</i>) and non-understory species (e.g., Yellow-margined Flycatcher—<i>Tolmomyias assimilis</i>) were captured less frequently, and continued sampling will likely yield longer longevity records for these species in particular. Finally, 11 of our longevity records were lower than those reported in other studies of shorter duration (Appendix S1: Table S1). In particular, studies of color-banded populations that have tracked individuals over long time periods (e.g., Tarwater & Kelley, <span>2020</span>; Willis, <span>1983</span>) have yielded longer longevity records than our community-level mark-recapture study, likely due to greater sampling effort and higher detection probability of color-banded individuals. Thus, our data suggest that even 40+ years of mark-recapture effort may not be sufficient to produce accurate longevity estimates for certain Neotropical forest bird species.</p><p>Future work that endeavors to address questions about how longevity varies geographically and assess its ecological relevance as a life history trait may require large-scale collaborative banding schemes and long-term population studies of banded birds in tropical regions to obtain broader geographical coverage and more accurate longevity estimates. Currently, the differences in spatial coverage and number of captures between our dataset (one site, thousands of captures) and large-scale banding schemes such as the BBL in North America and EURING in Europe (thousands of sites, millions of captures) span orders of magnitude, making direct comparison challenging. One potential way to test for geographical differences in longevity is standardized comparisons from single-site banding operations (e.g., banding sites included in the Institute for Bird Populations' Monitoring Avian Productivity and Survivorship: https://www.birdpop.org/pages/maps.php) across similar time periods. These long-term datasets are particularly difficult to compile in tropics, however, where biodiversity is greatest but sampling effort still lags behind that of temperate systems (Collen et al., <span>2008</span>). For example, Scholer et al. (<span>2018</span>) found that avian longevity estimates of Neotropical landbirds were highly correlated with study duration, highlighting the need for longer-term studies and the systemic challenges to amassing such longevity datasets in the tropics. Therefore, despite the moderately high correlation (<i>R</i><sup>2</sup> = 0.5) we found between survival and longevity, we concur with the conclusions of Krementz et al. (<span>1989</span>), who found that model-based estimates of survival were preferable to observed maximum lifespan statistics for use in comparative life history studies.</p><p>The authors declare no conflicts of interest.</p>\",\"PeriodicalId\":11484,\"journal\":{\"name\":\"Ecology\",\"volume\":\"105 9\",\"pages\":\"\"},\"PeriodicalIF\":4.4000,\"publicationDate\":\"2024-07-24\",\"publicationTypes\":\"Journal Article\",\"fieldsOfStudy\":null,\"isOpenAccess\":false,\"openAccessPdf\":\"https://onlinelibrary.wiley.com/doi/epdf/10.1002/ecy.4386\",\"citationCount\":\"0\",\"resultStr\":null,\"platform\":\"Semanticscholar\",\"paperid\":null,\"PeriodicalName\":\"Ecology\",\"FirstCategoryId\":\"93\",\"ListUrlMain\":\"https://onlinelibrary.wiley.com/doi/10.1002/ecy.4386\",\"RegionNum\":2,\"RegionCategory\":\"环境科学与生态学\",\"ArticlePicture\":[],\"TitleCN\":null,\"AbstractTextCN\":null,\"PMCID\":null,\"EPubDate\":\"\",\"PubModel\":\"\",\"JCR\":\"Q1\",\"JCRName\":\"ECOLOGY\",\"Score\":null,\"Total\":0}","platform":"Semanticscholar","paperid":null,"PeriodicalName":"Ecology","FirstCategoryId":"93","ListUrlMain":"https://onlinelibrary.wiley.com/doi/10.1002/ecy.4386","RegionNum":2,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
Long-term monitoring reveals the long lifespans of Neotropical forest landbirds
Longevity, or maximum lifespan, is a trait that has fascinated ecologists, evolutionary biologists, and geneticists for centuries and has featured prominently in the development of life history, aging, and senescence theories (Lankester, 1870; Nussey et al., 2013; Stearns, 1992). Longevity has been associated with a variety of other life history and morphological traits such as brain and body size (Lindstedt & Calder, 1981; Stearns, 1992), energy metabolism (Furness & Speakman, 2008; White & Seymour, 2004), hibernation strategy (Turbill et al., 2011; Wilkinson & South, 2002), and antioxidant capacity (Vágási et al., 2019). Meaningful longevity data, however, necessitate studies of marked individuals over extended periods, a requirement that can be challenging for long-lived vertebrate species. Long-term mark-recapture efforts provide some of the best opportunities for obtaining longevity estimates from free-living populations of animals. For example, among birds, continent-wide banding programs in North America (i.e., U.S.G.S. Bird Banding Laboratory [BBL]: https://www.pwrc.usgs.gov/bbl/longevity/longevity_main.cfm) and Europe (i.e., EURING: https://euring.org/data-and-codes/longevity-list) comprise decades of individual-based monitoring that have yielded longevity estimates for many temperate-zone species. By contrast, longevity data from tropical birds are few and restricted in temporal scope due to the logistical and financial challenges of maintaining long-term monitoring programs in the tropics (Collen et al., 2008; Proença et al., 2017). As of 2023, fewer than 250 longevity records of Neotropical landbirds are available in the published literature (reviewed in Scholer et al., 2018), and only two studies spanned ≥20 years (Arendt, 2006; Woodworth et al., 1999).
How long do Neotropical forest landbirds actually live? To address this question, we used a 45-year (1977–2022) mark-recapture dataset from central Panama—one of the longest of its kind in the Neotropics—to derive longevity records for 47 species. We sampled birds on the 100-ha Limbo Plot on Pipeline Road in the 22,000-ha Soberanía National Park (9.15° N, 79.73° W). The Limbo Plot is classified as tropical lowland moist forest and has been the focus of numerous previous studies of tropical bird ecology (e.g., Brawn et al., 2017; Karr, 1976; Karr & Freemark, 1983; Pollock et al., 2020, 2022; Tarwater et al., 2011; see Robinson et al., 2000 for a more detailed discussion of the Limbo Plot). Netting and banding protocols are described in detail in Pollock et al. (2022). Briefly, we mist-netted and banded birds twice per year (dry and wet season) at two 2-ha sites embedded in the larger 100-ha Limbo Plot. At each netting site, nets were set at ground level and kept open for approximately 600 total net-hours each sampling period. To estimate longevity, we compiled capture histories of more than 15,000 individuals of 150 species, indexed by unique band numbers. We excluded species with ≤20 total captures over the 45-year study period, as they were unlikely to use the understory stratum sampled by mist-nets or too locally rare to yield accurate longevity estimates. We then derived longevity records for 198 individuals of 47 tropical bird species in 17 families (henRyPollock, 2024 at https://doi.org/10.5281/zenodo.11106481). We report the minimum longevity (i.e., the elapsed time between initial banding and last recapture, sensu Scholer et al., 2018) of the longest-lived individual for each of the 47 species (Appendix S1: Table S1).
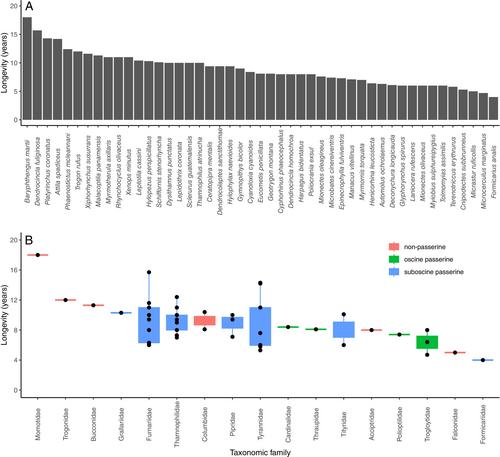
The oldest individual longevity record was an 18-year-old Rufous Motmot (Baryphthengus martii; Figure 1A). Other notably long-lived individuals included a Plain-brown Woodcreeper (Dendrocincla fuliginosa; 15.7 years; Figure 1B), two Golden-crowned Spadebills (Platyrinchus coronatus; 14.0 and 14.3 years; Figure 1C), two Bright-rumped Attilas (Attila spadiceus; 12.0 and 14.2 years; Figure 1D), an Ocellated Antbird (Phaenostictus mcleannani; 12.4 years; Figure 1E), and a Black-throated Trogon (Trogon rufus; 12.0 years; Figure 1F). Of the 47 longevity estimates included in the dataset (Figure 2A), 37 (79%) were the longest on record and 27 (57%) were first records for a given species, respectively (Appendix S1: Table S1). Most species (34/47; 72%) had a longevity of ≥7 years, 29 (62%) had a longevity of ≥8 years, and 18 (38%) had a longevity of ≥10 years (Appendix S1: Figure S1).
Of the 17 families included in the dataset, 14 (82%) had at least one species with a longevity record of ≥8 years and 10 (59%) had at least one species with a longevity record of ≥10 years (Figure 2B). Families with high longevities included the ovenbirds (Furnariidae: 6 of 9 species with ≥8 years longevity) and the antbirds (Thamnophilidae: all nine species ≥7 years; 7 of 9 species with ≥8 years longevity). Flycatchers (Tyrannidae) had a bimodal distribution, with very high longevities for some species (e.g., >14 years; Bright-rumped Attila and Golden-crowned Spadebill) and relatively low longevities (<7 years) for most other species despite similar recapture rates (Appendix S1: Table S1; henRyPollock, 2024). We tested for and found no significant differences in longevity between oscine passerines, suboscine passerines, and non-passerines (generalized linear model [GLM]: F2,44 = 2.0, p = 0.15, R2 = 0.08; Appendix S1: Figure S2).
We also tested for potential relationships between species' longevities and ecological traits including body mass, diet (i.e., frugivore, insectivore, omnivore, carnivore), and sociality (i.e., non-flocker, facultative flocker, obligate flocker, ant-follower, lekking species). We found no significant associations between longevity and body mass (GLM: F1,45 = 0.84, p = 0.36, R2 = 0.02; Appendix S1: Figure S3), diet (GLM: F4,42 = 1.0, p = 0.42, R2 = 0.09), or sociality (GLM: F4,42 = 1.10, p = 0.37, R2 = 0.09). Individuals captured more frequently or with higher abundances were expected to have higher longevity estimates, but we found no relationship between longevity and the number of times an individual was captured (GLM: F1,196 = 2.0, p = 0.16, R2 = 0.01; Appendix S1: Figure S4) or the total number of captures per species (GLM: F1,39 = 0.82, p = 0.37, R2 = 0.02). Finally, longevity has been questioned for its value as a life history trait versus, for example, annual survival rate (Krementz et al., 1989). Therefore, we tested for and found a moderately high positive correlation for 18 species from our study site for which estimates of both annual survival rates (derived from Brawn et al., 2017) and longevities were available (GLM: F1,16 = 3.05, p < 0.001, R2 = 0.50; Appendix S1: Figure S5).
The duration and depth of our study (45 years, >80,000 captures) provided an unprecedented window into the lives of Neotropical forest birds and revealed that many Neotropical species have considerably longer lifespans than previously documented in the scientific literature. We report the oldest longevity records to date for ~80% of all species and ~65% of all families sampled, encompassing a broad range of phylogenetic and ecological variation. Surprisingly, and in contrast with other studies (e.g., Furness & Speakman, 2008; Lindstedt & Calder, 1981), we found no correlation between body mass and longevity, perhaps because our focal species comprised a relatively narrow range of body masses (7–219 g; Appendix S1: Table S1) relative to other datasets. In fact, some of the smallest-bodied species (<15 g) at our study site were also the longest-lived, including Plain Xenops (Xenops minutus—11.0 years), White-flanked Antwren (Myrmotherula axillaris—11.0 years) and Golden-crowned Spadebill (Platyrinchus coronatus—14.3 years). Finally, non-passerines are thought to have longer lifespans (Lindstedt & Calder, 1976), but we found no evidence of differences in longevity between passerines and non-passerines, which we expect was due to having few non-passerines (n = 6 species) in our dataset, all of which had relatively small body sizes (42–219 g). Overall, our results reveal the long lifespans of tropical landbirds and reinforce the importance of long-term studies for obtaining these types of data (Magurran et al., 2010).
Nevertheless, several lines of evidence suggest that our longevity records are still likely biased underestimates of true longevity. First, the limited spatial extent of our study (two 2-ha plots) means that many individuals that were not recaptured could still have been persisting undetected outside of the study plots. Second, longevity was not correlated with the average number of recaptures of the sampled species, and nearly 60% of longevity records were based on individuals recaptured five or fewer times. For example, species with large home-ranges like army-ant-following birds (e.g., Ocellated Antbird—Phaenostictus mcleannani) and non-understory species (e.g., Yellow-margined Flycatcher—Tolmomyias assimilis) were captured less frequently, and continued sampling will likely yield longer longevity records for these species in particular. Finally, 11 of our longevity records were lower than those reported in other studies of shorter duration (Appendix S1: Table S1). In particular, studies of color-banded populations that have tracked individuals over long time periods (e.g., Tarwater & Kelley, 2020; Willis, 1983) have yielded longer longevity records than our community-level mark-recapture study, likely due to greater sampling effort and higher detection probability of color-banded individuals. Thus, our data suggest that even 40+ years of mark-recapture effort may not be sufficient to produce accurate longevity estimates for certain Neotropical forest bird species.
Future work that endeavors to address questions about how longevity varies geographically and assess its ecological relevance as a life history trait may require large-scale collaborative banding schemes and long-term population studies of banded birds in tropical regions to obtain broader geographical coverage and more accurate longevity estimates. Currently, the differences in spatial coverage and number of captures between our dataset (one site, thousands of captures) and large-scale banding schemes such as the BBL in North America and EURING in Europe (thousands of sites, millions of captures) span orders of magnitude, making direct comparison challenging. One potential way to test for geographical differences in longevity is standardized comparisons from single-site banding operations (e.g., banding sites included in the Institute for Bird Populations' Monitoring Avian Productivity and Survivorship: https://www.birdpop.org/pages/maps.php) across similar time periods. These long-term datasets are particularly difficult to compile in tropics, however, where biodiversity is greatest but sampling effort still lags behind that of temperate systems (Collen et al., 2008). For example, Scholer et al. (2018) found that avian longevity estimates of Neotropical landbirds were highly correlated with study duration, highlighting the need for longer-term studies and the systemic challenges to amassing such longevity datasets in the tropics. Therefore, despite the moderately high correlation (R2 = 0.5) we found between survival and longevity, we concur with the conclusions of Krementz et al. (1989), who found that model-based estimates of survival were preferable to observed maximum lifespan statistics for use in comparative life history studies.
期刊介绍:
Ecology publishes articles that report on the basic elements of ecological research. Emphasis is placed on concise, clear articles documenting important ecological phenomena. The journal publishes a broad array of research that includes a rapidly expanding envelope of subject matter, techniques, approaches, and concepts: paleoecology through present-day phenomena; evolutionary, population, physiological, community, and ecosystem ecology, as well as biogeochemistry; inclusive of descriptive, comparative, experimental, mathematical, statistical, and interdisciplinary approaches.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


