Margaret W. Simon, Michael Barfield, Nicholas Kortessis, S. Luke Flory, Keith Clay, Robert D. Holt
{"title":"Fast-growing annual plants drive disease spillover in multi-host communities","authors":"Margaret W. Simon, Michael Barfield, Nicholas Kortessis, S. Luke Flory, Keith Clay, Robert D. Holt","doi":"10.1111/1365-2745.70002","DOIUrl":null,"url":null,"abstract":"<h2>1 INTRODUCTION</h2>\n<p>Emerging infectious plant diseases threaten natural communities, agricultural crops (Nazarov et al., <span>2020</span>) and food security (Brooks et al., <span>2021</span>; Fones et al., <span>2020</span>; Ristaino et al., <span>2021</span>; Savary & Willocquet, <span>2020</span>), and synergize with other global change factors to exacerbate already declining plant communities (Anderson et al., <span>2004</span>). For example, when plant invaders accumulate pathogens that spill over onto native competitors, emerging infectious disease can magnify negative invader impacts on native flora (Kendig et al., <span>2022</span>). A major challenge in understanding and controlling emerging disease outbreaks and spillover is predicting which populations are most at risk. Because the potential for outbreaks can be modulated by other community members (Power & Mitchell, <span>2004</span>), monitoring for disease emergence and spillover requires surveilling not only a population of concern (the focal host), but also the full community within which it is imbedded.</p>\n<p>Disease spillover requires movement of infectious propagules from an individual of one host species (a reservoir host) to an individual of a second species (a focal host). Such a process can take one of two main forms: (i) there is an initial spillover event (i.e. propagule movement between two individuals of different species), with no further transmission needed to sustain disease in the focal host, or (ii) disease following the initial spillover event is augmented by further spillover events (Lloyd-Smith et al., <span>2009</span>; Power & Mitchell, <span>2004</span>). In the zoonotic and wildlife disease literature (e.g. see Lloyd-Smith et al., <span>2009</span>), these two spillover types have qualitatively different implications for disease establishment as understood by <span data-altimg=\"/cms/asset/23ee2621-fddf-4766-9930-f9a5529eae1d/jec70002-math-0001.png\"></span><mjx-container ctxtmenu_counter=\"0\" ctxtmenu_oldtabindex=\"1\" role=\"application\" sre-explorer- style=\"position: relative;\" tabindex=\"0\"><mjx-lazy aria-hidden=\"true\" data-mjx-lazy=\"0\"></mjx-lazy><mjx-assistive-mml display=\"inline\" unselectable=\"on\"><math data-semantic-=\"\" data-semantic-role=\"unknown\" data-semantic-speech=\"\" data-semantic-type=\"empty\" xmlns=\"http://www.w3.org/1998/Math/MathML\"></math></mjx-assistive-mml></mjx-container> (the average number of new infections, resulting from an initial infection event). A single spillover event cannot lead to disease establishment unless <span data-altimg=\"/cms/asset/4d62cea5-4cd7-4ace-82ce-a320962a4432/jec70002-math-0002.png\"></span><mjx-container ctxtmenu_counter=\"1\" ctxtmenu_oldtabindex=\"1\" role=\"application\" sre-explorer- style=\"position: relative;\" tabindex=\"0\"><mjx-lazy aria-hidden=\"true\" data-mjx-lazy=\"1\"></mjx-lazy><mjx-assistive-mml display=\"inline\" unselectable=\"on\"><math data-semantic-=\"\" data-semantic-role=\"unknown\" data-semantic-speech=\"\" data-semantic-type=\"empty\" xmlns=\"http://www.w3.org/1998/Math/MathML\"></math></mjx-assistive-mml></mjx-container> of the focal host exceeds unity. When <span data-altimg=\"/cms/asset/0f43cd1d-142c-4325-962c-63204b3183d3/jec70002-math-0003.png\"></span><mjx-container ctxtmenu_counter=\"2\" ctxtmenu_oldtabindex=\"1\" role=\"application\" sre-explorer- style=\"position: relative;\" tabindex=\"0\"><mjx-lazy aria-hidden=\"true\" data-mjx-lazy=\"2\"></mjx-lazy><mjx-assistive-mml display=\"inline\" unselectable=\"on\"><math data-semantic-=\"\" data-semantic-role=\"unknown\" data-semantic-speech=\"\" data-semantic-type=\"empty\" xmlns=\"http://www.w3.org/1998/Math/MathML\"></math></mjx-assistive-mml></mjx-container> < 1 in the focal host, a single initial spillover event is not sufficient for disease persistence. However, the disease may nonetheless persist if there is continual pathogen transmission from further spillover events. In traditional SIR systems, <span data-altimg=\"/cms/asset/ca508840-2a80-4ff1-8450-08382cee36c7/jec70002-math-0004.png\"></span><mjx-container ctxtmenu_counter=\"3\" ctxtmenu_oldtabindex=\"1\" role=\"application\" sre-explorer- style=\"position: relative;\" tabindex=\"0\"><mjx-lazy aria-hidden=\"true\" data-mjx-lazy=\"3\"></mjx-lazy><mjx-assistive-mml display=\"inline\" unselectable=\"on\"><math data-semantic-=\"\" data-semantic-role=\"unknown\" data-semantic-speech=\"\" data-semantic-type=\"empty\" xmlns=\"http://www.w3.org/1998/Math/MathML\"></math></mjx-assistive-mml></mjx-container> is defined at the beginning of an outbreak when all individuals are susceptible and generally scales directly with disease transmission rate, abundance of the susceptible class and the duration of host infection (Keeling & Rohani, <span>2008</span>).</p>\n<p>Translations of this framework to plant disease spillover remain a challenge (Thompson & Brooks-Pollock, <span>2019</span>), in major part due to important differences in how plant pathogens spread compared with animal pathogens. For example, plants often experience localized disease. Host plants, by nature of their modularity, can be treated conceptually as an entity containing both infected and noninfected (i.e. susceptible) parts, rather than each individual being either entirely infected or susceptible as in many epidemiological models. The potential for an individual to transmit disease (or become infected) could scale with its amount of infected (or susceptible) parts. We propose that individual host plant size could act like host density in animal infectious disease systems, where higher host densities usually increase disease transmission rates (Keeling & Rohani, <span>2008</span>; Power & Mitchell, <span>2004</span>).</p>\n<p>In addition to their modularity, plants are largely sessile, meaning that initial infection is driven by pathogen, rather than host, movement as typically envisioned in animal models. Pathogen propagules can move among individual hosts via wind or rain splash (Bourouiba, <span>2021</span>; Mukherjee et al., <span>2021</span>), or vectors, such as aphids. Here, our focus will be on pathogen movement by abiotic modes and self-dispersal, such as ballistic dispersal. In the case of many ascomycete and basidiomycete pathogens, pathogen spores disperse by launching from fruiting bodies (Trail, <span>2007</span>) or are self-propelled in dew droplets (Mukherjee et al., <span>2021</span>). Therefore, larger host plants with more surface area should provide more opportunities for pathogen propagules to contact the host. Plant size can change by orders of magnitude very quickly (Cunniffe et al., <span>2015</span>), the timescale of which, for some plants (e.g. annuals) is comparable to that over which disease progression occurs, suggesting that dynamical feedbacks between plant growth and pathogen infection and spread could occur.</p>\n<p>These differences between plant-disease dynamics and typical animal-based models might help explain current uncertainties around the occurrence of species richness on disease in plant communities. One hypothesis is that species diversity amplifies disease in communities, while a second hypothesis poses that species diversity dilutes disease (Keesing et al., <span>2006</span>, <span>2010</span>; Ostfeld & Keesing, <span>2012</span>, and references in Perkins & Rohr, <span>2020</span>). We posit that relative differences in size may play an important, but unrecognized, mechanistic role in the amplification/dilution debate in plant communities.</p>\n<p>The goal of this paper was to develop an understanding of the effect of individual plant growth on the potential for, and effects of, pathogen spillover in plant communities. In predator–prey apparent competition, prey species with high intrinsic growth rates sustain greater predator abundances, which then suppress other, slower-growing, prey species in the community (Holt & Lawton, <span>1994</span>). We hypothesize that a similar phenomenon could be occurring among plant species with different individual growth rates that share a common pathogen, the idea being that the rate of growth in the size of a plant host individual could serve a similar purpose to a prey's population growth rate. We explore this possibility by developing a host–pathogen model incorporating the changing size structure of, and degree of infection in, annual plant host species within a growing season. Specifically, we aim to determine how species differences in plant growth rates alter (i) the potential for spillover from a reservoir population, (ii) the impacts of spillover and (iii) the amplification or dilution of disease in communities comprised of two alternative (non-competing) host populations. We first explore how disease proceeds within a single plant in isolation. We then examine disease spillover from a reservoir population onto a focal population, without spillback onto the reservoir. (The focal population in this case can be thought of as a novel host in which the pathogen has evolved the ability to infect, but has not yet evolved the ability to spillback onto the reservoir). Finally, we model disease dynamics in a community of two host species that reciprocally share a pathogen, with approximately equal interspecific transmission of the pathogen between the two hosts.</p>","PeriodicalId":191,"journal":{"name":"Journal of Ecology","volume":"63 1","pages":""},"PeriodicalIF":5.3000,"publicationDate":"2025-02-10","publicationTypes":"Journal Article","fieldsOfStudy":null,"isOpenAccess":false,"openAccessPdf":"","citationCount":"0","resultStr":null,"platform":"Semanticscholar","paperid":null,"PeriodicalName":"Journal of Ecology","FirstCategoryId":"93","ListUrlMain":"https://doi.org/10.1111/1365-2745.70002","RegionNum":1,"RegionCategory":"环境科学与生态学","ArticlePicture":[],"TitleCN":null,"AbstractTextCN":null,"PMCID":null,"EPubDate":"","PubModel":"","JCR":"Q1","JCRName":"ECOLOGY","Score":null,"Total":0}
引用次数: 0
Abstract
1 INTRODUCTION
Emerging infectious plant diseases threaten natural communities, agricultural crops (Nazarov et al., 2020) and food security (Brooks et al., 2021; Fones et al., 2020; Ristaino et al., 2021; Savary & Willocquet, 2020), and synergize with other global change factors to exacerbate already declining plant communities (Anderson et al., 2004). For example, when plant invaders accumulate pathogens that spill over onto native competitors, emerging infectious disease can magnify negative invader impacts on native flora (Kendig et al., 2022). A major challenge in understanding and controlling emerging disease outbreaks and spillover is predicting which populations are most at risk. Because the potential for outbreaks can be modulated by other community members (Power & Mitchell, 2004), monitoring for disease emergence and spillover requires surveilling not only a population of concern (the focal host), but also the full community within which it is imbedded.
Disease spillover requires movement of infectious propagules from an individual of one host species (a reservoir host) to an individual of a second species (a focal host). Such a process can take one of two main forms: (i) there is an initial spillover event (i.e. propagule movement between two individuals of different species), with no further transmission needed to sustain disease in the focal host, or (ii) disease following the initial spillover event is augmented by further spillover events (Lloyd-Smith et al., 2009; Power & Mitchell, 2004). In the zoonotic and wildlife disease literature (e.g. see Lloyd-Smith et al., 2009), these two spillover types have qualitatively different implications for disease establishment as understood by (the average number of new infections, resulting from an initial infection event). A single spillover event cannot lead to disease establishment unless of the focal host exceeds unity. When < 1 in the focal host, a single initial spillover event is not sufficient for disease persistence. However, the disease may nonetheless persist if there is continual pathogen transmission from further spillover events. In traditional SIR systems, is defined at the beginning of an outbreak when all individuals are susceptible and generally scales directly with disease transmission rate, abundance of the susceptible class and the duration of host infection (Keeling & Rohani, 2008).
Translations of this framework to plant disease spillover remain a challenge (Thompson & Brooks-Pollock, 2019), in major part due to important differences in how plant pathogens spread compared with animal pathogens. For example, plants often experience localized disease. Host plants, by nature of their modularity, can be treated conceptually as an entity containing both infected and noninfected (i.e. susceptible) parts, rather than each individual being either entirely infected or susceptible as in many epidemiological models. The potential for an individual to transmit disease (or become infected) could scale with its amount of infected (or susceptible) parts. We propose that individual host plant size could act like host density in animal infectious disease systems, where higher host densities usually increase disease transmission rates (Keeling & Rohani, 2008; Power & Mitchell, 2004).
In addition to their modularity, plants are largely sessile, meaning that initial infection is driven by pathogen, rather than host, movement as typically envisioned in animal models. Pathogen propagules can move among individual hosts via wind or rain splash (Bourouiba, 2021; Mukherjee et al., 2021), or vectors, such as aphids. Here, our focus will be on pathogen movement by abiotic modes and self-dispersal, such as ballistic dispersal. In the case of many ascomycete and basidiomycete pathogens, pathogen spores disperse by launching from fruiting bodies (Trail, 2007) or are self-propelled in dew droplets (Mukherjee et al., 2021). Therefore, larger host plants with more surface area should provide more opportunities for pathogen propagules to contact the host. Plant size can change by orders of magnitude very quickly (Cunniffe et al., 2015), the timescale of which, for some plants (e.g. annuals) is comparable to that over which disease progression occurs, suggesting that dynamical feedbacks between plant growth and pathogen infection and spread could occur.
These differences between plant-disease dynamics and typical animal-based models might help explain current uncertainties around the occurrence of species richness on disease in plant communities. One hypothesis is that species diversity amplifies disease in communities, while a second hypothesis poses that species diversity dilutes disease (Keesing et al., 2006, 2010; Ostfeld & Keesing, 2012, and references in Perkins & Rohr, 2020). We posit that relative differences in size may play an important, but unrecognized, mechanistic role in the amplification/dilution debate in plant communities.
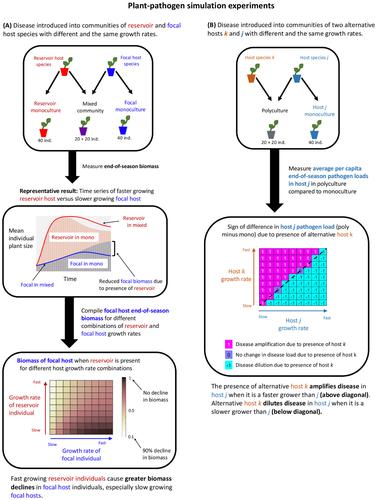
The goal of this paper was to develop an understanding of the effect of individual plant growth on the potential for, and effects of, pathogen spillover in plant communities. In predator–prey apparent competition, prey species with high intrinsic growth rates sustain greater predator abundances, which then suppress other, slower-growing, prey species in the community (Holt & Lawton, 1994). We hypothesize that a similar phenomenon could be occurring among plant species with different individual growth rates that share a common pathogen, the idea being that the rate of growth in the size of a plant host individual could serve a similar purpose to a prey's population growth rate. We explore this possibility by developing a host–pathogen model incorporating the changing size structure of, and degree of infection in, annual plant host species within a growing season. Specifically, we aim to determine how species differences in plant growth rates alter (i) the potential for spillover from a reservoir population, (ii) the impacts of spillover and (iii) the amplification or dilution of disease in communities comprised of two alternative (non-competing) host populations. We first explore how disease proceeds within a single plant in isolation. We then examine disease spillover from a reservoir population onto a focal population, without spillback onto the reservoir. (The focal population in this case can be thought of as a novel host in which the pathogen has evolved the ability to infect, but has not yet evolved the ability to spillback onto the reservoir). Finally, we model disease dynamics in a community of two host species that reciprocally share a pathogen, with approximately equal interspecific transmission of the pathogen between the two hosts.
期刊介绍:
Journal of Ecology publishes original research papers on all aspects of the ecology of plants (including algae), in both aquatic and terrestrial ecosystems. We do not publish papers concerned solely with cultivated plants and agricultural ecosystems. Studies of plant communities, populations or individual species are accepted, as well as studies of the interactions between plants and animals, fungi or bacteria, providing they focus on the ecology of the plants.
We aim to bring important work using any ecological approach (including molecular techniques) to a wide international audience and therefore only publish papers with strong and ecological messages that advance our understanding of ecological principles.

 求助内容:
求助内容: 应助结果提醒方式:
应助结果提醒方式:


